
Interactions between plant nutrients: A deep understanding

On this page:
Plant nutrient interactions occur when one nutrient influences the uptake, availability, or utilization of another within the soil–plant system. These relationships can be synergistic, improving nutrient use efficiency and crop yield, or antagonistic, limiting uptake. Understanding these interactions helps growers optimize fertilization strategies and maintain balanced plant nutrition.
Introduction
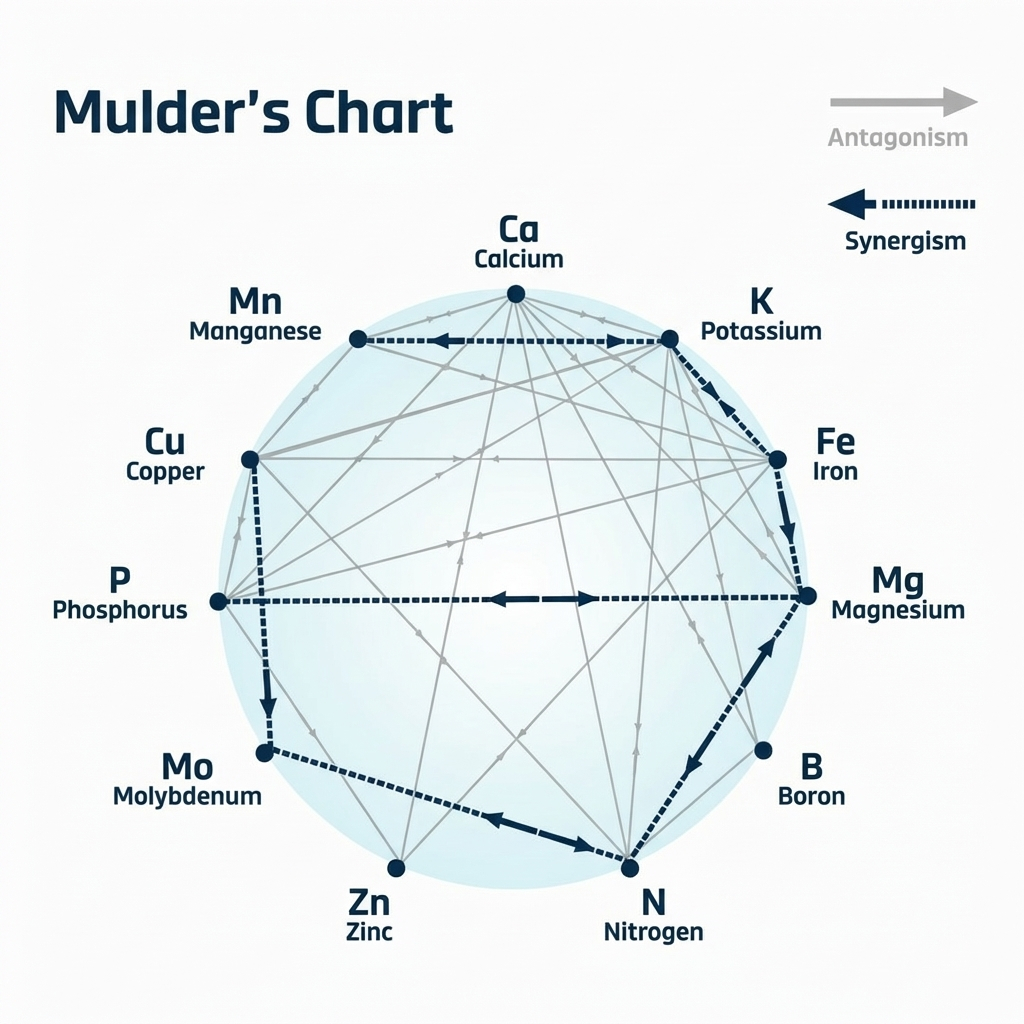
Plants require a balance of macronutrients (e.g., N, P, K, Ca, S, Mg) and micronutrients (Fe, Zn, Cu, Mn, B, Mo, etc.) for growth. (Kumar et al., 2021). Nutrient “interactions” occur when one element influences the uptake or use of another. (Rietra et al., 2017). These interactions can be synergistic or antagonistic. For example, applying N and K together often boosts yield more than either alone (synergy). In contrast, an excess of one cation (like Zn2+ or Mg2+) can inhibit the uptake of another (antagonism). In general, most macronutrient pairs (N–P, N–K, etc.) show synergistic effects on yield and uptake, whereas many divalent cations (Ca, Mg, Fe, Mn, Zn, Cu) commonly antagonize each other because they compete for the same transport pathways (Rietra et al., 2017).
Nutrient Use Efficiency (NUE) in Crop Production
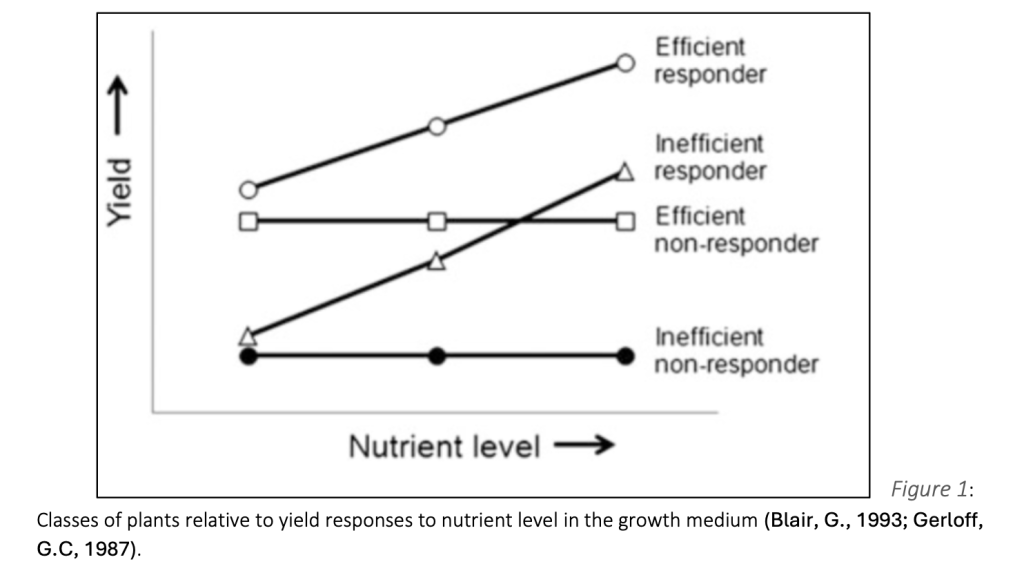
Plants’ nutrient use efficiency (NUE) is greatly affected by physiological and genetic components, and their effect on plants’ ability to uptake and utilize nutrients under different environmental conditions (Nieves-Cordones et al., 2020). NUE determination can be beneficial for distinguishing plant species, genotypes, and cultivars by their ability to uptake and assimilate nutrients for maximal dry matter and yield production. NUE is composed from three efficiency mechanisms:
- Uptake efficiency: absorption from the soil, influx kinetics and influx rate into the roots, radial transport (based on root parameters per length or weight), uptake is also correlated to particular nutrient amounts that are present in soil or were applied,
- Incorporation efficiency: transport to the shoot and leaves (based on shoot parameters), and
- Utilization efficiency: based on remobilization and the whole plant parameters. Plant NUE may be defined as the maximum economic yield or dry matter produced per unit of an applied nutrient or a unit of that nutrient taken up (Baligar, V.C & Fageria, N.K, 2015).
Nutrient interactions profoundly affect NUE. Synergistic fertilization, which provides complementary nutrients, usually increases overall NUE. For example, combining N with K will allow similar yields with less N fertilizer, effectively increasing N use efficiency (Rietra et al., 2017). In practice, balanced NPK (with micronutrients) means each applied unit of N, P, or K is more fully utilized in plant growth, and less is lost to leaching or denitrification. Conversely, a severe imbalance (e.g., very high N without K) can decrease NUE because the missing nutrient limits yield. Improving NUE in crops, therefore, relies on managing these interactions. Ensuring that all required elements are sufficiently available without being in gross excess allows plants to achieve high yields with less fertilizer.
For example, low sulfur (S) can cap protein synthesis even if N is ample, so co-applying S with N often boosts NUE.
Macronutrient Interactions (N, P, K)
Nitrogen (N), phosphorus (P), and potassium (K) often work together in plant metabolism, and their availability can affect each other’s uptake and use. For example, sufficient N improves P acquisition. Conversely, P deficiency can inhibit N assimilation. This N–P synergy means that crops supplied with both N and P usually produce higher yields than when given either nutrient alone. Similarly, N–K interactions are generally positive: applying both N and K fertilizers typically results in higher yields and better N-use efficiency compared to N alone (Rietra et al., 2017).
The interaction of K and N is historically well documented, with experiments started in 1852 at Rothamsted Station, UK (Ranade-Malvi, 2011). Some of the interactions that affect crop response to a nutrient like K are quality factors such as fertilizer form, method, application date, variety of crops, and more. The occurrence of such interactions may lead to changes in the ways of using K fertilizers. The most important interaction is with quantitative variables, such as the level of other nutrients applied, irrigation rate, plant spacing, etc. Among these, the interaction between K and other nutrients, particularly N, is the most important (Loue, 1980).
Another important function of K application could be to further improve N metabolism enzyme activity (Hu, W. et al., 2016; Zahoor et al., 2017). The significance of N-K interaction and how to manage it is increasing due to higher crop yield, increasing cropping intensity, and considerable K depletion in cultivated soils (Aulakh & Malhi, 2005).
Crops with high K requirements often show strong N-K interactions (Loue, A., 1980; Singh, M, 1992). Plants take up N either in cationic (NH4+) or in anionic (NO3-) form. This creates a unique option of anion–cation and cation–cation interactions with K. Most of the findings have shown that K does not compete with NH4+ for uptake, but increases NH4+ assimilation in the plants and prevents possible NH4+ toxicity (Aulakh & Malhi, 2005).
Mengel et al. (1976) determined that it is improbable for K to compete with NH4+ for selective binding sites during the uptake process. To read more specifically about N-K interactions, please refer to Perelman et al., 2021.
Synergistic and Complementary Nutrient Interactions in Plants
- N and P interactions: A recent long-term field experiment across multiple species found that often plants show co-limitation by N and P. This means giving both together produces a stronger growth response than either alone. Plant N and P acquisition processes are strongly interrelated, and combined N and P fertilization should be considered in fertilizer recommendations (Spohn, 2025).
- N and K interactions: a recent review on N–K interactions showed that adequate external K⁺ supply enhances uptake and translocation of nitrate (NO₃⁻), the main N form, from roots to shoots, because K⁺ often acts as a counter-ion to NO₃⁻ in xylem and phloem transport. Under K deficiency, NO₃⁻ translocation is impaired, reducing N use efficiency (Cao et al., 2025).
- P and K interactions: Liu et al., (2024) showed that in rice growing in hydroponics, sufficient P and K supply, vs. low supply, significantly enhanced root growth, root morphology, N absorption, N utilization efficiency, and photosynthetic N metabolism (enzymes, assimilation). This shows that P and K availability can indirectly boost N uptake and utilization. Long-term fertilization experiments showed that application of N, P, and K together, compared to single-nutrient or dual mixes, resulted in the highest K uptake and overall crop productivity (Arbačauskas et al., 2023).
N, P, and K often work in complementary ways. Successive or combined supply can lead to more efficient uptake and use of N, better biomass, root development, and yield, especially when plants are not already saturated by one nutrient. Balanced fertilization (all three) often outperforms single- or dual-nutrient fertilization.
Antagonistic and Limiting Nutrient Interactions in Plants
- Low soil K can limit N response: when applying large amounts of N when soil exchangeable K is poor, it gives poor NUE and limited yield response because, without enough K, the plant cannot sustain the increased water content, osmotic balance, and NO₃⁻ translocation required for N utilization (Perelman et al., 2021).
- High K supply can suppress other cations/nutrient balance: Some studies report that increased K may negatively impact uptake of other nutrients, especially divalent cations like Mg and Ca, which may indirectly impair overall nutrition or increase risk of
secondary nutrient deficiencies. For instance, in a hydroponic study of non-crop plants, increasing K significantly decreased uptake of NH₄⁺ and reduced accumulation of other cations (Ca, Mg) (Tzortzakis et al., 2025). - NPK ratios matter: over-supplying one may deplete others. In long-term studies, applying only N (without balanced P and K) resulted in lower soil K availability and reduced K uptake over time (Arbačauskas et al., 2023).
Physiological/mechanistic basis of NPK interactions
Understanding nutrient interactions at a mechanistic level helps explain why fertilizer imbalances cause poor efficiency or toxicity. Some key points:
- Ion co-transport and counter-ions: Especially for N and K, when plants take up nitrate (NO₃⁻),
K⁺ often acts as a counter-ion to maintain charge balance as nitrates are transported via
xylem/phloem. Adequate external K⁺ thus supports efficient NO₃⁻ uptake and translocation.
Under low K conditions, NO₃⁻ loading into the xylem is impaired, reducing N uptake and
utilization (Cao et al., 2025). - Root morphology and uptake surface: Adequate P and K encourage better root development
(e.g., longer roots, better root density), which enlarges the root surface area and improves
nutrient absorption capacity (for N, P, K). In a rice study, P+K improved root length, root
weight, and overall root health, which in turn improved N uptake and N use efficiency (Liu et
al., 2024). - Photosynthesis and assimilation enzymes: P and K availability influences leaf physiology:
insufficient P or K reduces chlorophyll content, photosystem II activity, and ATP synthesis,
which reduces photosynthesis; this lowers energy available for assimilation (e.g., nitrogen
assimilation into amino acids and proteins), thereby decreasing nitrogen-use efficiency (El-
Mageed et al., 2023; Liu et al., 2024). - Interlinked signaling and transport networks: Research in model plants shows that uptake and
signaling pathways for K and P (and N) are interconnected. For example, certain transporters
and regulatory proteins coordinate uptake of K and P, meaning that deficiency or abundance
in one can influence the regulation of the other (Fan et al., 2021).
Secondary Nutrients (Ca, Mg, S) and Their Interactions with Macronutrients
The secondary (meso) nutrients calcium (Ca), magnesium (Mg), and sulfur (S) also interact strongly with each other and with NPK. Chemically, K+, Ca2+, and Mg2+ are all cations taken up by root ion channels, so an excess of one often inhibits another. Experimental studies in hydroponics show that too much Ca inhibits Mg uptake, and too much K inhibits Ca uptake (Palani, 2019; Rietra et al., 2017).
Similarly, soil tests and agronomic experience report that high K fertilization reduces plant Mg availability, while excess Mg can make K less available (Xie et al., 2021).
Ca and Mg can antagonize each other; excess Ca also impedes Mg uptake. These cationic interactions occur because the nutrients share similar transporters and binding sites on root membranes (Jakobsen, 1993).
Nitrogen and sulfur interact through their roles in the synthesis of amino acids and proteins. S deficiency limits synthesis of cysteine and methionine, thus restraining N utilization in proteins; conversely, high N demand increases S uptake. (Mechanistic details of N–S co-regulation are complex and involve changes in amino acid pools and root signaling.) In brief, balanced N and S fertilization is known to improve protein yield and NUE in many crops.
Macronutrient–Micronutrient Interactions
Macros also affect micronutrient uptake. A classic case is P–Zn antagonism. Excessive soil P can induce Zn deficiency. A field study showed that the Zn concentration in wheat grain decreased as more P fertilizer was applied. High P tends to precipitate or immobilize Zn, especially in alkaline soils, so plants take up less Zn. Similarly, application of high N, especially as ammonium, can acidify the rhizosphere and transiently increase availability of Fe, Mn, and other cationic micros, whereas nitrate (alkalinizing) can decrease it (Rietra et al., 2017).
Many micronutrients are cations that compete with each other and with Ca/Mg. For example, Fe, Mn, Zn, Cu, and Ca all use overlapping transporters (ZIP, NRAMP, COPT families, etc.). Studies show that antagonism is common among these divalent cations (Rietra et al., 2017).
For instance, a combined Zn+Mg treatment in wheat produced a lower growth response than expected, implying an antagonistic effect. Likewise, excess Zn can down-regulate the iron transporter IRT1 in roots, causing Fe deficiency (Kumar et al., 2021).
In general, synergisms among Fe, Mn, Zn, Cu, Ca, or Mg rarely occur. Rietra et al., (2017) notes that most antagonistic interactions involve these cations, presumably because they compete for uptake mechanisms. Some synergistic crosstalk between macro- and micro-nutrients exists; for example, adequate N often enhances Fe and Zn uptake indirectly by promoting root growth and organic acid exudation. Adequate P can increase root and mycorrhizal development, sometimes improving micronutrient uptake. These positive effects are often conditional, for example, only when a micronutrient is not severely deficient. Overall, nutrient uptake in plants is regulated by complex molecular networks (transporters, chelators, signaling pathways, etc.), many of which respond to multiple nutrient cues. For example, the PHR1 transcription factor links N, P, S, Fe, and Zn homeostasis, and specific transporters (e.g., NRT1.1B-SPX4) tie N signaling to P uptake. Such mechanisms help plants coordinate
the acquisition of nutrients under varying conditions (Kumar et al., 2021).
Bibliography
- Arbačauskas, J., Vaišvila, Z. J., Staugaitis, G., Žičkienė, L., Masevičienė, A., & Šumskis, D. (2023). The Influence of Mineral NPK Fertiliser Rates on Potassium Dynamics in Soil: Data from a Long-Term Agricultural Plant
Fertilisation Experiment. Plants, 12(21), 3700. https://doi.org/10.3390/plants12213700 - Aulakh, M. S & Malhi, S. S. (2005). Interactions of nitrogen with other nutrients and water: Effect on crop yield
and quality, nutrient use efficiency, carbon sequestration, and environmental pollution. Advances in Agronomy,
.409–341 ,86 - Baligar, V.C & Fageria, N.K. (2015). Nutrient Use Efficiency in Plants: An Overview. In Nutrient Use Efficiency:
From Basics to Advances (pp. 1–14). Springer. - Blair, G. (1993). Nutrient efficiency—What do we really mean? In Genetic Aspects of Plant Mineral Nutrition.
Developments in Plant and Soil Sciences (Vol. 50). Springer. - Cao, W., Sun, H., Shao, C., Wang, Y., Zhu, J., Long, H., Geng, X., & Zhang, Y. (2025). Progress in the Study of Plant Nitrogen and Potassium Nutrition and Their Interaction Mechanisms. Horticulturae, 11(8), 930.
https://doi.org/10.3390/horticulturae11080930 - El-Mageed, T. A. A., Semida, W. M., Abdou, N. M., & El-Mageed, S. A. A. (2023). Coupling Effects of Potassium
Fertilization Rate and Application Time on Growth and Grain Yield of Wheat (Triticum aestivum L.) Plants Grown Under Cd-Contaminated Saline Soil. Journal of Soil Science and Plant Nutrition, 23(1), 1070–1084.
https://doi.org/10.1007/s42729-022-01104-3 - Fan, X., Zhou, X., Chen, H., Tang, M., & Xie, X. (2021). Cross-Talks Between Macro- and Micronutrient Uptake
and Signaling in Plants. Frontiers in Plant Science, 12, 663477. https://doi.org/10.3389/fpls.2021.663477 - Gerloff, G.C. (1987). Intact-plant screening for tolerance of nutrient-deficiency stress. Plant Soil, 99, 3–16.
https://doi.org/10.1007/BF02370149 - Hu, W., Zhao, W, Yang, J, Oosterhuis, D. M, Loka, D. A, & Zhou, Z. (2016). Relationship between potassium
fertilization and nitrogen metabolism in the leaf subtending the cotton (Gossypium hirsutum L.) boll during the
boll development stage. Plant Physiology and Biochemistry, 10, 113–123. - Jakobsen, S. T. (1993). Interaction between Plant Nutrients: III. Antagonism between Potassium, Magnesium and Calcium. Acta Agriculturae Scandinavica, Section B-Soil & Plant Science, 43(1), 1–5. https://doi.org/10.1080/09064719309410223
- Kumar, S., Kumar, S., & Mohapatra, T. (2021). Interaction Between Macro‐ and Micro-Nutrients in Plants.
Frontiers in Plant Science, 12, 665583. https://doi.org/10.3389/fpls.2021.665583 - Liu, Y., Gao, J., Zhong, M., Chen, L., & Zhang, W. (2024). Effects of Phosphorus and Potassium Supply on
Photosynthetic Nitrogen Metabolism, Nitrogen Absorption, and Nitrogen Utilization of Hydroponic Rice.
Agronomy, 14(8), 1726. https://doi.org/10.3390/agronomy14081726 - Loue, A. (1980). The interaction of potassium with other growth factors particularly with other nutrients. 1st
Congr. Int. Potash Inst (pp. 67–93). International Potash Institute. - Mengel, K, Viro, M, & Hehl, G. (1976). Effect of potassium on uptake and incorporation of ammonium-nitrogen
of rice. Plant and Soil, 44(3), 547–558. - Nieves-Cordones, M., Rubio, F., & Santa-María, G. E. (2020). Editorial: Nutrient Use-Efficiency in Plants: An
Integrative Approach. Frontiers in Plant Science, 11, 623976. https://doi.org/10.3389/fpls.2020.623976 - Palani, V. (2019). Synergistic and Antagonistic Interactions of Calcium with Other Nutrients in Soil and Plants.
SSRN Electronic Journal. https://doi.org/10.2139/ssrn.3503225 - Perelman, A., Imas, P., & Bansal, S. K. (2021). Role of Potassium for Improving Nutrient Use Efficiency in
Agriculture. In R. Bhatt, R. S. Meena, & A. Hossain (Eds.), Input Use Efficiency for Food and Environmental
Security (pp. 397–420). Springer Nature Singapore. https://doi.org/10.1007/978-981-16-5199-1_13 - Ranade-Malvi, U. (2011). Interaction of micronutrients with major nutrients with special reference to potassium.
- Rietra, R. P. J. J., Heinen, M., Dimkpa, C. O., & Bindraban, P. S. (2017). Effects of Nutrient Antagonism and
Synergism on Yield and Fertilizer Use Efficiency. Communications in Soil Science and Plant Analysis, 48(16),
1895–1920. https://doi.org/10.1080/00103624.2017.1407429 - Singh, M. (1992). The nitrogen–potassium interaction and its management. In Management of Nutrient
Interactions in Agriculture. Fertiliser Development and Consultation Organisation. - Spohn, M. (2025). Interactions of nitrogen and phosphorus in plant nutrition—Analysis of a 60-years old field
experiment. Plant and Soil, 510(1–2), 275–289. https://doi.org/10.1007/s11104-024-06920-3 - Tzortzakis, N., Neofytou, G., & Chrysargyris, A. (2025). Exploring the Effect of Nitrogen, Phosphorus and
Potassium Rates of the Nutrient Solution on the Growth and Quality of Common Sowthistle (Sonchus oleraceus
L.) Grown Hydroponically. Journal of Soil Science and Plant Nutrition, 25(3), 7946–7960.
https://doi.org/10.1007/s42729-025-02644-0 - Xie, K., Cakmak, I., Wang, S., Zhang, F., & Guo, S. (2021). Synergistic and antagonistic interactions between
potassium and magnesium in higher plants. The Crop Journal, 9(2), 249–256.
https://doi.org/10.1016/j.cj.2020.10.005 - Zahoor, R., Zhao, W., Abid, M., Dong, H., & Zhou, Z. (2017). Potassium application regulates nitrogen
metabolism and osmotic adjustment in cotton (Gossypium hirsutum L.) functional leaf under drought stress.
Journal of Plant Physiology, 215, 30–38. https://doi.org/10.1016/j.jplph.2017.05.001
