
Supplement to Basic Knowledge of Paddy Rice Cultivation
On the Roles of Secondary and Micro-nutrients and Crop Physiology by Growth Stage in Paddy Rice Cultivation

On this page:
This comprehensive guide expands on the fundamentals of paddy rice cultivation, exploring soil preparation, water management, nutrient balance, and growth stages essential for optimal yield. It connects traditional knowledge with modern agronomic practices, emphasizing how environmental conditions and nutrient availability affect rice physiology. A valuable reference for growers and agronomists alike, it offers practical insights for improving productivity, sustainability, and crop quality across diverse cultivation systems.
Japan, located in the eastern part of the Asian monsoon region, has a climate with abundant rainfall and lowland wetland soils (Aquepts, Alluvial soils) that are well suited for paddy rice cultivation. Rice farming has long been encouraged in the country. Improving rice yield has been a national policy, and universities and research institutions have actively conducted research on rice cultivation. From the 1950s to the 1980s, Japan became a global hub for rice cultivation research, during which a vast amount of knowledge were accumulated and agricultural techniques were developed. In particular, studies on the physiology and nutrition of paddy rice significantly contributed to the advancement of rice farming techniques.
Thanks to scientifically grounded agricultural practices—including fertilization methods, selective breeding, soil improvement, pest and disease control, and mechanization—yield-limiting factors were progressively overcome. By 1970, rice production had reached the point where Japan even had a surplus supply, both in terms of quantity and quality.
It has now been over half a century since this era of integrated efforts by the government and private sectors, and the environment surrounding rice cultivation has changed drastically. Researches on rice cultivation at universities and public institutions have largely disappeared. In recent years, there has been a growing demand to shift toward more sustainable paddy rice cultivation methods that are resilient to climate change and aligned with various needs, such as environmental conservation, reduced chemical fertilizer use, and organic agriculture.
Future technologies for rice cultivation will still rely on the application of past scientific knowledge. By supplementing the “Crop Nutrient Advice on Rice” with Japan’s accumulated expertise in rice traits and morphology, plant nutrition and physiology, fertilization methods, and diagnostic techniques—recognized globally—it is hoped that this knowledge will contribute, even in a small way, to the future success of those who engaged in rice farming.
Physiological Roles of Secondary and Micronutrients
Sulfur (S)
Sulfur is one of the essential elements and a component of proteins. The sulfur and nitrogen content in proteins is roughly equivalent, and both are necessary for building the plant body during early growth stages. Sulfur is absorbed by roots in the form of sulfate ions (SO₄²⁻), then transported directly to the stems and leaves, where it is reduced and assimilated in the chloroplasts to form sulfur compounds.
When sulfur is deficient, protein synthesis declines, resulting in stunted rice growth and halted tillering. Sulfur deficiency typically appears in the early growth stages. Symptoms include smaller leaves with pale yellow-green coloration and yellowing of the lower leaves. The number of tillers is reduced, and root development is also weakened.
In general, sulfur deficiency tends to occur in paddy soils with low natural sulfur supply, such as mineral soils low in organic matter. Deficiency is more likely to manifest when available sulfur levels are below 20 ppm. Additionally, when soil reduction progresses and excess iron and manganese are produced, the solubility of sulfur decreases, leading to sulfur deficiency.
In recent years, even paddy fields that have long relied on sulfate-free fertilizers have begun to show signs of sulfur deficiency due to declining soil sulfur concentrations.
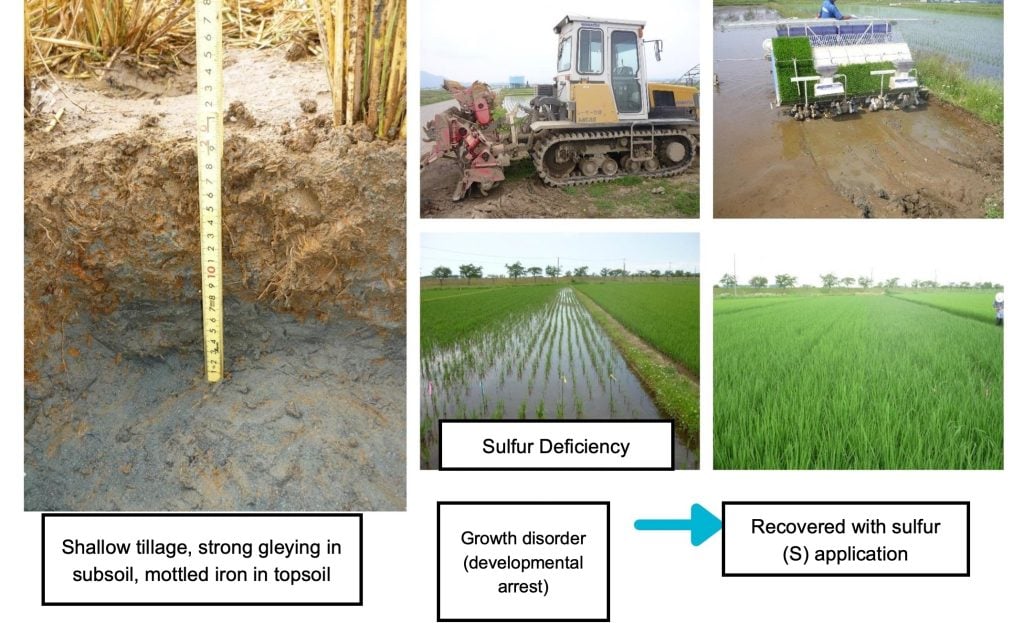
Sulfur Deficiency in Paddy Rice (on Strongly Gley Soils). Photos courtesy of Dr. Shin Hidaka (Japan)
Manganese (Mn)
Manganese is one of the essential micronutrients for paddy rice. Under flooded conditions, manganese is dissolved in the soil solution in its divalent form (Mn²⁺) and absorbed by the roots as Mn²⁺ ions. It is transported to the leaves via transpiration and is primarily found in the chloroplasts.
As a transition metal, manganese plays a vital role in physiological functions such as oxygen evolution during photosynthesis and the activation of various enzymes. It is an essential component of the electron transport system, making it critical for plant metabolism.
Generally, under typical paddy field conditions, manganese becomes available, and deficiency is rare. However, in aging paddy fields or sandy soils, manganese can leach out easily, leading to an absolute shortage of plant-available manganese in the topsoil. Fields with easily reducible manganese levels below 10 ppm (optimal levels are 50–80 ppm or more) are considered to have low manganese supply capacity.
Deficiencies in potassium, silica, manganese, and magnesium during the mid to late stages of rice growth can trigger Brown Spot (caused by filamentous fungi). This disease is frequently observed in sandy soils where fertilizers are prone to leaching or in “Akiochi” paddy fields where root rot is common.

Leaching of Iron and Manganese into Subsoil in Paddy Fields.Photos courtesy of Dr. Shin Hidaka (Japan)
Silicon (Si)
Although silicon is not classified as an essential element, it is a beneficial element that significantly influences the growth of paddy rice. Rice plants absorb around 100 kg of silicic acid per 10 ares. While large quantities of silicate compounds exist in the soil, most are insoluble, and the supply capacity of silicic acid in paddy soils is generally low.
Sandy soils derived from granite have particularly low silicic acid availability, and soils with high phosphate absorption coefficients tend to adsorb more silicic acid, reducing its availability to plants. The primary source of available silicic acid for paddy rice is organic matter in the soil, such as rice straw and compost, followed by silicate-based soil amendments and irrigation water.
When the concentration of plant-available silicic acid in paddy soil is 15 mg/100g or less, the application of silicate fertilizers is expected to be effective. Rice actively absorbs water-soluble orthosilicic acid (H₄SiO₄) through root respiration. It is then transported through the xylem via transpiration flow to aboveground tissues, where it polymerizes and deposits as silica (SiO₂) in the epidermal cells of leaves, leaf blades, and the outer layer of husks.
In the epidermal tissues of leaves, silica gel accumulates between the cell wall and the cuticle layer. This strengthens the physical structure of stems and leaves, enhancing lodging resistance, disease resistance (by preventing fungal invasion), improving light interception, and boosting photosynthesis—ultimately contributing greatly to yield and quality improvement.
When silicon is deficient, the rice plants become weak overall, panicle length is shortened, and yield is reduced. Additionally, brown discoloration of grains (such as brown rice defects) may occur, severely degrading grain quality.
Thus, the usefulness of silicon in rice is expressed through its accumulation in epidermal cells, which helps avoid water stress in leaves and enhances the structural integrity of the plant. Silicon is also effective in mitigating heat stress, such as reducing the incidence of white immature grains (mature grains are translucent).
Iron (Fe)
In soil, iron exists in two oxidation states: ferrous iron (Fe²⁺) and ferric iron (Fe³⁺). In aerobic soils, most of the iron is present as poorly soluble Fe³⁺. However, under flooded (anaerobic) conditions, Fe³⁺ is reduced to Fe²⁺, which is more soluble and readily absorbed by rice plants.
In general, paddy fields under flooded conditions have significantly high concentrations of Fe²⁺ in the soil solution. To prevent excessive uptake, rice roots possess an iron exclusion mechanism that suppresses iron absorption. However, in fields with poor drainage, excess iron can lead to iron toxicity (known as Akagare, or “red leaf disease”), which manifests as reddish-brown spots on leaves and withering from the tips and edges.
One effective countermeasure is the use of potassium fertilizers, which enhance the root’s ability to exclude iron and thereby alleviate the symptoms.
When soil pH is high, iron precipitates as ferric hydroxide (Fe(OH)₃), a poorly soluble form, reducing iron availability to plants. Additionally, excess levels of heavy metals such as copper (Cu), manganese (Mn), and zinc (Zn) can trigger iron deficiency (chlorosis) due to antagonistic interactions, characterized by yellowing or whitening of young leaves and localized brown spotting.
Compared to other crops, paddy rice has a relatively high demand for iron and will actively absorb it when concentrations are low. Within the plant, iron exists mainly as Fe²⁺, some of which binds with specific proteins to form iron-protein complexes. While iron is not a structural component of chlorophyll, it participates in redox reactions in both Fe²⁺ and Fe³⁺ states and is critical in:
- The electron transport system of photosynthesis
- Energy metabolism
- Activation of various enzymes
In flooded soils, free ferric oxide (Fe oxides) also plays an important role by protecting roots from toxic substances like hydrogen sulfide and organic acids, helping to prevent root rot.
In aging paddy fields or sandy soils, free iron oxides tend to leach into the subsoil, lowering iron levels in the topsoil. Ideally, the content of free iron oxide in paddy soil should be at least 0.8%. In iron-deficient fields, applying iron-rich materials such as converter slag (200 kg/10a) helps maintain root vigor and can improve rice yields.
Boron (B)
In soil, boron exists as boric acid (H₃BO₃) bonded with oxygen, or as its salts. In the soil solution, it appears either as molecular boric acid or as the B(OH)₄⁻ ion. The form of boron varies depending on soil pH:
- In acidic soils (pH < 7), H₃BO₃ predominates and is considered plant-available boron.
- In neutral to alkaline soils (pH ≥ 7), H₃BO₃ concentrations decrease, and B(OH)₄⁻ becomes more prevalent.
Boron is absorbed by plants in the form of BO₃³⁻ and transported via transpiration to the stems and leaves.
Although rice has a relatively low demand for boron and deficiencies are rare, high-pH soils reduce the availability of plant-available boron, making deficiency more likely. Boron is essential for maintaining cell wall structure and also plays a role in the transport of sugars produced by photosynthesis.
Boron deficiency affects actively dividing tissues. In rice, typical symptoms include:
- Stunted early growth
- Reduced number of panicles and grains per panicle
- Increase in sterile grains (empty husks)
- Poor heading due to inhibited pollen tube elongation
The optimal range of plant-available boron (hot-water-extractable) in soils is narrow, between 0.5 and 1.0 mg/kg. Deficiencies often occur below 0.3 mg/kg.
Soils that are sandy or sandy loam, with low cation exchange capacity or poor in humus, tend to have weak boron adsorption and are prone to boron deficiency.
On the other hand, boron toxicity in rice can occur if:
- Plant-available boron exceeds 1.0 mg/kg in general plants
- Water-soluble boron exceeds 10 mg/kg in rice
Excess boron in rice is often caused by:
- Overuse of boron-rich fertilizers
- Contaminated wastewater from plating or paint industries
Symptoms of excess include:
- Browning of leaf tips
- White spots are forming on the leaves
- Progressive leaf bleaching
Countermeasure: Apply heavy irrigation to leach out excess boron from the soil.
Zinc (Zn)
In soil, zinc exists in several forms:
- As crystalline compounds in soil minerals,
- As organic forms bound to plant residues and humic substances,
- And as exchangeable or water-soluble forms on clay particle surfaces.
However, a significant portion is present as insoluble forms such as zinc sulfide (ZnS), zinc hydroxide (Zn(OH)₂), or zinc carbonate (ZnCO₃).
The availability of zinc in soil is influenced by several factors:
- Soil pH: Zinc is more soluble in acidic soils, while it becomes unavailable in neutral to alkaline conditions.
- Excessive phosphate application can form insoluble zinc compounds.
- Adsorption by oxides of iron, aluminum, or manganese suppresses plant absorption.
- Field conditions: In reclaimed or lowland paddies, reduced conditions can lead to the formation of zinc sulfide, resulting in zinc deficiency.
- On the other hand, industrial wastewater from zinc mines or factories can cause zinc toxicity.
- Zinc-deficient soils also include serpentine or peridotite-derived Paleozoic soils.
Rice absorbs zinc primarily as divalent Zn²⁺ ions, though in high-pH soils, it may also be taken up as ZnOH⁺. Once absorbed, free zinc is transported through the xylem to growing tissues in stems and leaves, where it binds with proteins to form complexes.
Physiological roles of zinc:
- It plays a role in protein synthesis and in the biosynthesis of plant growth hormones such as auxin.
- Deficiency symptoms include:
- Shortened stems and leaf blades
- Reduced tillering
- “Akagare” symptoms (a type of bronzing caused by physiological stress)
- Weak growth and lodging
- Increased number of sterile grains
Rice is highly sensitive to zinc deficiency, and symptoms can occur when soil zinc concentrations fall below 15 mg/kg. Acid sulfate soils are particularly prone to zinc deficiency due to inherently low zinc content.
As an emergency measure, zinc sulfate application (surface broadcast) has been shown to improve conditions.
On the other hand, zinc toxicity from pollution (e.g., industrial or mining runoff) appears during early growth stages, with symptoms such as:
- Interveinal chlorosis on young leaves
- More prominently, root damage: excessive branching or “lion tail” root malformation
Three Types of “Akagare” (Bronzing) Disorders in Rice
- Akagare Type I:
Caused by iron toxicity
– Occurs in poorly drained sandy soils, black mud soils, and acid sulfate soils.
– Known as “Bronzing.” - Akagare Type II:
Caused by zinc deficiency
– Due to an absolute shortage of zinc, high-pH soils, or poor drainage. - Akagare Type III:
Caused by iodine excess
– Results from the conversion of upland to paddy fields, where decomposition of soil organic matter and reduction conditions lead to excessive absorption of soluble iodide (I⁻).
Molybdenum (Mo)
Molybdenum occurs in minerals primarily as molybdenite (MoS₂) and in nature as oxidized, poorly soluble molybdenum trioxide (MoO₃). In general, molybdenum has low solubility, and its concentration in soil ranges from 0.1 to 6.0 mg/kg, varying greatly depending on soil type.
Molybdenum availability in soil is highly pH-dependent:
- At pH 5–6 and above, it dissolves as the molybdate ion (MoO₄²⁻) and becomes more water-soluble.
- Below that, it exists as HMoO₄⁻, often adsorbed by iron (Fe) and aluminum (Al) oxides or bound to organic matter.
- In more strongly acidic soils, it becomes unavailable as molecular molybdic acid (H₂MoO₄).
Thus, deficiency tends to occur in acidic soils, while toxicity may occur in alkaline soils.
Rice absorbs molybdenum in the form of molybdate ions (MoO₄²⁻), which are transported through the xylem. Among all essential elements, molybdenum has the lowest required quantity in plants.
Physiologically, it is:
- A component of nitrate reductase, an enzyme involved in nitrate assimilation,
- It is also involved in phosphate metabolism, photosynthesis, and respiration.
Deficiency symptoms include:
- Nitrate accumulation in plant tissues,
- And reduced photosynthetic activity.
Generally, plants show deficiency symptoms when molybdenum content is below 0.1 ppm. In rice straw, molybdenum levels are typically low, ranging from 0.2 to 0.7 ppm in most cases.
Crop Physiology by Growth Stage
Seed
A seed develops from a fertilized endosperm. In rice, the seed is referred to as the “momi” (grain with husk) and is enclosed in two layers: the lemma (outer husk) and palea (inner husk). Inside are the embryo and the endosperm, and collectively (including the husk), this structure is called the seed.
- The embryo consists of the plumule (young shoot) and radicle (young root)—the part that germinates.
- The endosperm is filled primarily with starch granules, which store nutrients necessary for embryo development.
- The outermost layer of the endosperm, called the aleurone layer, is rich in proteins and fats and plays a critical role in supplying nutrients to the embryo during germination.
The seed is a vital part of the rice life cycle. After fertilization and maturation, it separates from the parent plant, settles in a new location, and grows into a new rice plant, carrying the genetic information of its parent.
Rice actively absorbs silicic acid from the soil even after the heading stage. The husk (momi) can contain over 10% silica, which serves a critical defensive function, protecting the seed from heat, drought, pathogens, and pests—a specialized adaptation to wetland environments unique to rice.
Immediately after harvest, seeds exhibit dormancy, a temporary inhibition of germination. This phenomenon ensures the seed doesn’t germinate prematurely. The aging process (loss of viability) begins as soon as the seed matures on the parent plant.
The two most critical factors influencing seed dormancy and lifespan are moisture content and temperature. Therefore, seed storage should begin immediately after harvest.
For seeds to be used the following year, post-threshing and cleaning, the moisture content should be reduced to 11–14%. The seeds should then be sealed in airtight containers and stored in a cool, dark, indoor environment.
To select high-germination-quality seed rice, a saltwater flotation test is used:
- Place seeds in saltwater with a specific gravity of 1.06.
- Collect the sunken grains (which are well-filled with nutrients in the endosperm).
- Rinse off the salt thoroughly with fresh water.
Seedling
To germinate, dry seeds first require water. Once the embryo is fully hydrated, germination begins. The endosperm near the embryo absorbs water as well, and the decomposition products of stored nutrients are supplied to the growing shoot and root after germination—this process is called “seed soaking.”
The duration of seed soaking depends on water temperature, with a total accumulated temperature of 100°C (water temperature × days) being the standard. Germination also requires a large amount of oxygen, so during soaking, the water should be refreshed regularly to maintain adequate oxygen levels. Oxygen deficiency during soaking causes abnormal elongation of the shoot (plumule) and inhibits root (radicle) development.
When using a germination chamber with an oxygen supply, germination from soaking can be achieved within one day.
Once the seed has absorbed enough water and the embryo is slightly swollen (“pigeon chest” stage), it is sown into seedling soil, soft and well-aerated. The seeds are then placed in a growth chamber with temperature control (30–32°C) to promote uniform germination.
- The first leaf emerges in about 2 days in the chamber.
- After that, the seedlings are transferred to 20–25°C conditions for greening, followed by hardening outdoors under natural sunlight and ambient temperature.
Up to the emergence of the second leaf, the seedling relies on nutrients from the endosperm. Starting from the third leaf stage (“weaning stage”), when the endosperm nutrients are depleted, the seedling begins to absorb water and nutrients through its roots and perform photosynthesis, becoming nutritionally independent (autotrophic).
The period up to the fourth leaf stage—known as the tillering initiation stage—is critical. Sudden temperature spikes, deep water (causing oxygen deficiency and root damage), or excess nitrogen in the soil can lead to excessive elongation of leaf sheaths, resulting in weak, leggy growth and suppressed lower tillering.
Since tillers formed during the seedling stage tend to become productive panicles, it is important to avoid overgrowth and oxygen deficiency to produce healthy seedlings in a short time.
In mechanized transplanting systems using seedling trays, rice seedlings are classified by leaf age:
- Young seedlings :3–3.5 leaf stage
- Intermediate seedlings 4–5.5 leaf stage
Leaves
Rice leaves consist of the leaf blade and leaf sheath, with auricles (leaf ears) and a ligule attached.
- The leaf blade is the primary site for photosynthesis and transpiration, and it plays a key role in distributing nutrients throughout the plant. Most of its carbohydrates are in the form of hemicellulose, contributing to cell wall formation. Starch content is low, but soluble sugar concentration is high, allowing the leaf to function as a transport organ for assimilates.
- The leaf sheath wraps around the stem, providing protection and support, and temporarily stores assimilates from the blade.
- The auricles are located at the junction of the blade and sheath, are broad and thickened, and help support the blade against wind and physical strain.
- The ligule, a thin membrane at the top of the sheath near the stem, prevents water from entering between the leaf sheath and stem.
The main stem typically produces 14–15 leaves, which grow alternately on opposite sides of the lower nodes. Leaf growth is greatly influenced by temperature, and leaf age (leaf stage) is an important indicator of the rice plant’s developmental stage and guides cultivation management.
Different leaf stages serve distinct physiological roles:
- 3rd–4th leaf stage: Supports root development by sending carbohydrates to roots (helps seedling establishment).
- 4th–8th leaf stage: Critical for tiller formation.
- 8th–12th leaf stage: Determines panicle size and grain number.
- 12th to final leaf (flag leaf, ~15th leaf): Important for grain filling (ripening).
Key cultivation practices tied to leaf stages:
- Mid-season drainage is typically done around the 7th–8th leaf stage.
- Panicle fertilizer is applied around the 11th–12th leaf stage, when young panicle development (~1.5–2 mm long) is confirmed.
To optimize topdressing timing, farmers can use:
- Iodine–starch reaction (described below), or
- Asparagine test (also detailed below).
The photosynthetic capability of leaves varies depending on their position:
- The top 3–4 leaves are most photosynthetically active and serve as the physiologically active center leaves.
- These leaves support the development of new organs and overall plant growth.
- During tillering, assimilated starch supports leaf growth; at young panicle formation, it supports the panicle; and after flowering, it contributes to grain filling.
As growth progresses:
- The active center leaves shift upward.
- Assimilates from older (lower) leaves are more likely to be redirected to the roots.
- The flag leaf remains an active center until the end and is critical for grain ripening.
- The upper 3–4 leaves, including the flag leaf, are essential for grain filling.
As older leaves lose nitrogen from their chlorophyll, photosynthesis declines, and nutrients are transferred to developing organs. Yellowing indicates the end of their functional life.
Field Diagnostic Techniques
Iodine–Starch Reaction Test
- Crush the 3rd leaf sheath down from the top, fully expanded leaf on the main stem.
- Soak for 1 minute in a diluted iodine-potassium iodide solution:
- Dissolve 2 g potassium iodide (KI) in 100 mL of water.
- Add 1 g iodine (I₂) and mix well. Store in a brown bottle.
- Measure the total sheath length and stained area.
- If the average staining rate (from 4–8 samples) is ≥55%, standard topdressing is appropriate.
- If ≤50%, postpone or skip topdressing.
Asparagine Test
- Asparagine presence correlates with high internal nitrogen; it disappears as nitrogen decreases.
- Serves as a panicle fertilizer indicator.
- Procedure:
- Soak the top and bottom leaves of the main culm in 70% ethanol.
- Grind the leaves in a mortar.
- Leave overnight in a cool, dark place.
- Evaporate and concentrate the supernatant.
- Apply to chromatography paper, run a second development with a solvent, then stain with ninhydrin.
- A gray spot indicates asparagine presence.
Note: Due to its complexity, the asparagine test is rarely used in the field.
Culm
The stem of rice is called the culm, which is composed of nodes and internodes as its basic structural units and is wrapped in leaf sheaths. The rice culm is usually not visible from the outside, except for the internodes that support the panicle. The internodes are arranged vertically at the base of the culm, overlapping one another. When the plant enters the reproductive growth stage, elongation begins from the lower internodes, and this elongation ends about 2–3 days after heading and flowering. The internodes are numbered from the top as the 1st to the 6th internode, and those below the 6th generally do not elongate.
Each node contains meristematic cells that produce new organs according to the rice plant’s growth, including leaves, tillers, roots, and panicles. The functions of the culm include:
- Support: Providing structural support for the rice plant.
- Transport: Conducting water and nutrients absorbed by the roots and photosynthetic products from the leaves to each organ.
- Storage: Temporarily storing photosynthates (mainly starch) synthesized in the leaves.
Nitrogen nutrition has a sensitive effect on the growth of each organ. If additional nitrogen is applied around 40 days before heading, or if nitrogen is abundant 20–30 days before heading, the 5th and 6th internodes tend to elongate, increasing the risk of lodging (plant collapse). A safe timing for panicle fertilization is when the internode length is 3.5 cm or less, and the young panicle is 1.5–2 mm long (young panicle formation stage).
Photosynthates in the leaves are converted to sucrose, transported, and stored as starch in the leaf sheath. When culm elongation begins, starch accumulates in the culm. Once panicle ripening begins, this starch immediately moves to the panicle. Although the starch storage in the culm is temporary, it plays an important role as a reservoir.
During the milky ripening stage, starch in the culm drastically decreases, and the transport of sugars necessary for root vitality also declines. If this coincides with a warm, dry wind event, the entire plant is at high risk of green wilting, which results in the occurrence of whiteheads (panicles that fail to fill).
Tiller (Tillering)
Tillers are newly grown stems that emerge as buds from the nodes at the base of the leaf sheath of the main stem. The stem and leaves are a collective body of tiller units, and each tiller consists of organs such as leaves, culms, roots, and panicles. When the leaves of the main stem emerge, a new tiller develops regularly from the node located below the third leaf. Once the main stem reaches its final leaf stage, all tiller leaves also reach their final leaf stage.
The development of tillers varies depending on planting density, number of transplanted seedlings, temperature, and nutrient conditions. Normally, the first tiller emerges after establishment when the fifth leaf of the main stem appears. Tillers that grow directly from the main stem are called primary tillers, and from their nodes, secondary tillers grow. These further branch into tertiary tillers, and so on. In general, primary and secondary tillers are dominant, while higher-order tillers (tertiary and above) are less productive. The higher the node from which a tiller emerges, the more likely it is to become a non-productive tiller.
Environmental stress—such as late transplanting, delayed root establishment, poor nutrient status during the tillering period, or light competition—often leads to growth stagnation or death of the tillers, resulting in non-productive tillers (those that do not form panicles).
In general, the required number of stems must be established within 30 days after transplanting. Rice tillering includes several stages:
- Maximum tillering stage: when the number of tillers peaks,
- Tillering peak period: about two weeks prior to the maximum tillering stage,
- Effective tiller ratio: the proportion of productive stems (that form panicles) out of the maximum tiller count.
A single rice plant typically produces about 20 tillers. If the number of plants per hill increases, competition causes suppression (dormancy) of lower node tillers and increases weak stems from higher nodes, which often do not produce panicles. A similar effect occurs with excessive nitrogen fertilization.
The number of tillers is influenced by the nitrogen content of the stems. Applying large amounts of nitrogen as base or supplemental fertilizer increases tiller formation but also leads to weak stems and a higher proportion of non-productive tillers. Since 60–70% of the nitrogen absorbed by rice comes from soil fertility, highly fertile paddy fields—especially those that undergo mid-season drainage—can lead to lodging (plant collapse).
In general, recommended base fertilizer amounts are based on local experience. In recent years, various slow-release fertilizers have become available. It is important to understand the nitrogen release type for optimal application.
Roots
The rice root system consists of a single seminal root that emerges from the seed and crown roots that develop from the basal nodes of the stem and leaves. Lateral roots branch out from these, forming a fibrous root system. Roots are considered part of a developmental unit along with leaves, nodes, tiller buds, and internodes.
Root development is closely related to leaf emergence. Crown roots (primary roots) from a given node grow in conjunction with the leaf three nodes above it. After germination, water and nutrient uptake are primarily handled by the seminal root. Starting from the 4-leaf stage, crown roots begin to take over this role, while the seminal root gradually loses its function.
Root elongation and nutrient/water uptake require carbohydrate supply from the aboveground parts, primarily from starch stored in the leaf sheath. Among the upper leaves, the third leaf from the top—called the active center leaf—has the highest starch accumulation in its sheath and is vascularly connected to the node just below it, from which vigorous crown roots emerge. As the plant grows, the active center leaf shifts upward, and so does the root-emitting node. In this way, root development progresses from lower nodes to upper nodes.
During the establishment phase, roots grow downward. As the plant develops, roots begin spreading laterally. Around the young plant’s formation stage, roots that emerge grow horizontally near the soil surface and form a root mat, made of primary roots arranged like a mesh.
The strongest rooting and elongation are observed during the peak tillering stage when photosynthesis is most active. As the plant transitions to the reproductive growth phase, root vitality and absorption of water and nutrients increasingly depend on photosynthates from the lower leaves. After heading, carbohydrates begin flowing predominantly to the panicle, reducing sugar concentration in the roots. This causes a drop in osmotic pressure within root cells and leads to a decline in water uptake.
By the heading stage, the root system is fully developed, consisting of various types:
- Old roots (primary roots)
- New roots and branching roots (secondary and tertiary roots)
- Deep roots
- Shallow roots
Each type plays a crucial role. Before heading, older roots contain a high concentration of sugars—key for energy—and provide much of the water and nutrients needed for shoot growth. New roots not only absorb nutrients and water but also possess strong oxidative capacity, which improves the rhizosphere environment and supports the absorption ability of older roots.
Deep roots are important for absorbing elements like iron, manganese, and silicon, which can easily leach in paddy fields. Shallow roots (root mats) continue to absorb water and nutrients during the ripening period, helping maintain the photosynthetic capacity of upper leaves and contributing to grain filling.
Regarding oxidation capacity, rice roots have well-developed aerenchyma (air-conducting tissue) that supplies oxygen from the leaves. This enables root respiration and nutrient absorption even under flooded, anaerobic conditions.
After heading and flowering, the photosynthetic activity of the plant becomes especially critical for grain production. Since leaf photosynthesis depends on moisture content, maintaining root vitality is essential. Continuous (saturated) irrigation is more effective than intermittent irrigation because it helps preserve the flow of starch from the lower leaves to the roots.
Saturated Irrigation (Saturated Water Management):
A method where the roots are kept in a state where they can always absorb water. Water is added when the field surface begins to dry (when footprints or furrows still retain water), and then allowed to drain naturally. This cycle is repeated.
Intermittent Irrigation:
Water is added only after the water in the footprints has completely disappeared. This method is typically used from mid-season drainage through the young panicle formation stage.
Heading and Flowering
The timing of heading varies significantly depending on the variety’s characteristics (such as sensitivity to light and temperature), temperature, day length, and region. This section describes the physiological traits of rice related to heading.
Heading refers to the process where the developing young panicle, enclosed in the leaf sheath, grows and is pushed upward as the internode at its base (called the neck node) elongates. It then emerges from the ligule of the flag leaf. The beginning of this emergence is called the start of heading; when about 40–50% of the tillers have headed, it is referred to as heading stage; and when more than 80% have headed, it is called full heading.
The main stem produces the first panicle, followed by those of the primary and secondary tillers. A rice panicle consists of the neck node, primary rachis branches, secondary rachis branches, and spikelets (grains).
The formation of the panicle begins around 32 days before heading, when the node that becomes the neck node is formed (neck node differentiation stage). Around 29 days before heading (spikelet differentiation stage), the primary branches develop from the neck node, followed by the secondary branches. After the secondary branches are formed, spikelets begin to develop on each branch.
Spikelets serve as carbohydrate storage organs and are composed morphologically of the hull (husk) and brown rice (grain). The rachis branches function as conduction tissues, transporting nutrients.
The spikelet formation period is approximately 25 days before heading. At this stage, anthers, stigmas, filaments, and endosperm begin to develop, and the spikelets enter the meiosis stage, around 12 days before heading. This period—from panicle initiation to meiosis—is highly sensitive to environmental conditions. If temperatures drop below 17°C or if there is a shortage of nitrogen or carbohydrates, the already formed branches and spikelets may degenerate, reducing the number of grains.
In paddy fields where fresh organic matter or high C/N ratio organic matter has been applied, organic acids generated under reducing conditions (Eh below -200 mV) can inhibit branch differentiation and lead to abnormal panicle development (called straight head disorder). This is a critical period in determining the number of grains per panicle.
In general, high temperatures promote heading, while low temperatures delay it. The optimal temperature for flowering is around 30°C, and flowering typically occurs between 10:00 and 12:00 on sunny days, lasting about 1.5 to 2 hours. On cloudy days, flowering is delayed.
At flowering, water demand in the panicle is high. If the panicle dries out due to strong winds or foehn phenomena, sterile grains can result. Additionally, if temperatures drop below 17°C or exceed 35°C during flowering, sterility disorders may occur.
About 7–10 days after flowering, the endosperm tissues are fully formed, and starch accumulation begins.
Grain Filling (Ripening)
The process by which the rice grain develops and enlarges after heading and flowering is called grain filling, and the period from this point until full maturity is known as the grain filling stage.
During this stage, the vascular transport system of each organ becomes highly active, and many of the components that make up the stems and leaves are redirected and transferred to the panicle. Although starch accumulation in the seed occurs over a short period, the transfer begins from the upper rachis branches and proceeds downward. Within the endosperm, accumulation starts at the center and progresses vertically, gradually increasing in thickness and fullness.
Approximately 20–40% of the starch found in the grains comes from stored starch in the leaf sheaths and culms formed before heading, while about 70% originates from photosynthesis that occurs after heading. Therefore, post-heading photosynthesis is extremely important for grain production.
The amount of starch in the grain peaks around 20–25 days after flowering, at which point the grains are considered physiologically mature. By 40–50 days after heading, grain filling is complete (dough stage).
Around 20 days after heading, internal stored starch used for energy—such as for respiration and water/nutrient uptake—decreases, and aging of the roots, stems, and leaves begins. However, roughly 70% of the starch still needed for grain filling must come from continued photosynthesis during this period. Thus, maintaining the vigor of the active leaves, including the flag leaf, is crucial.
To ensure adequate photosynthesis after heading, it’s important to avoid excessive vegetative growth and to properly manage the number of spikelets per panicle. This requires balanced cultivation management that harmonizes seedlings, leaves, culms, and tillers.
Water management becomes especially important after heading, including the use of saturated irrigation. One notable high-temperature issue during grain filling is the formation of chalky immature grains (white kernels). These occur frequently when the average temperature during the 20 days after heading exceeds 26–27°C.
The direct cause of chalky grains is high temperature during grain filling, which is closely related to the starch accumulation process in the endosperm. Starch is deposited in the following order: center, peripheral region, belly side, back side, and base of the endosperm. The specific area that turns chalky can indicate when the stress occurred.
- Milky-white grains at the center often result from excessive spikelet numbers, which reduce starch allocation per grain.
- Backside or base chalkiness increases when nitrogen is deficient during grain filling, reducing starch accumulation and causing gaps during deposition.
- Dead grains (fully white grains) occur when early drainage during the grain filling stage leads to water stress, reducing photosynthesis in the peripheral regions.
Regarding water and nitrogen management:
- Early drainage under nitrogen-deficient conditions can lead to cracked brown rice.
- Cadmium in the soil becomes more soluble under oxidative conditions. Early drainage during this period can significantly raise cadmium concentration in brown rice, so careful management is required.
Yield Components
Rice yield is determined by four main components:
- Number of panicles (per area)
- Number of grains per panicle
- Ripening ratio
- Grain weight
These four components function independently during the yield formation process but are interrelated and collectively determine the final yield.
- Panicle number is expressed per unit area by multiplying the number of panicles per plant by planting density. Since panicle number is determined by the maximum tillering stage, cultivation management from the seedling stage through tillering is crucial.
To ensure the panicle number:- Use healthy seedlings (short leaf sheaths)
- Plant shallowly (to encourage tillering from lower nodes)
- Avoid excessive nitrogen during fertilization
- Practice proper water management: Once the desired number of effective tillers (thick stems) is secured, start midseason drainage. The ideal timing is when about 80% of the target panicle number has been achieved.
- The number of grains per panicle is counted for many panicles and represented as an average. This is determined between the neck node differentiation stage (about 32 days before heading) and the end of meiosis (about 7 days before heading).
- Branch development is affected by environmental conditions around spikelet differentiation (about 24 days before heading).
- The final grain number is especially influenced by environmental factors during the panicle initiation to meiosis period, including light availability and nutrient status, which impact the formation and degeneration of branches and spikelets.
- Ripening ratio is the percentage of total spikelets that develop into filled, mature grains. It is measured by placing grains in saltwater (specific gravity 1.06) and calculating the ratio of sunken grains to total grains.
A low ripening ratio can result from:- Low temperatures during flowering (below 20°C)
- Strong winds
- Excess moisture (prolonged rain)
- High temperatures and dryness
Other factors include insufficient starch accumulation before heading, reduced photosynthetic activity after heading, excessive grain number, poor leaf posture (especially of the top 2–3 leaves), and a low grain-to-straw ratio.
The grain-to-straw ratio represents the balance between dry matter produced during the reproductive phase versus the vegetative phase. A higher ratio indicates stronger dry matter production during grain filling.
The ripening ratio is most sensitive during the meiosis stage through 15 days after heading, as this period determines photosynthate availability for grain development.
- Grain weight is expressed as the weight of 1,000 brown rice grains.
To calculate the estimated yield of polished brown rice per unit area:
Panicle number × Grains per panicle = Total grains per area
Total grains × Ripening ratio = Filled grains per area
Filled grains × Average grain weight = Final yield
Since calculated yield may differ from actual field results, average values for each component are typically used in yield estimation.
High-Yield Rice Cultivation Techniques
The previous sections, which have quoted from the book below, introduced the traits, organ development, and functions of rice. Although each topic was presented in a fragmented and limited manner, the content reflects findings clarified by many experts in Japan.
The challenge of achieving high rice yields begins with seedling production. Key points include:
- Producing healthy seedlings by ensuring proper oxygen supply during germination and seedling growth,
- Preventing overseeding and etiolation, and carefully managing temperature and water,
- Developing seedlings with strong root systems.
The rice plant is largely composed of organic matter, primarily proteins and carbohydrates. Proteins are mainly produced during the vegetative growth phase, from transplanting to neck node differentiation, and are used for tiller development. This stage is also when the straw portion of the grain-to-straw ratio is established.
To prevent excessive vegetative growth before heading, planting density should be adjusted based on expected light capture (light posture). Proper fertilization and water management should ensure the right number of effective tillers, maintain good light exposure, and prevent lodging.
Restricting nitrogen uptake during this period improves light posture and forms the foundation for increasing the grain filling ratio. A midseason drainage (drying) period helps improve root health.
The reproductive growth stage—from panicle initiation to heading—is centered on carbohydrate metabolism, and it is the most important time for determining grain number and accumulating starch in the leaf sheaths and culms.
During this stage, monitor:
- Leaf color and density,
- Young panicle length (1.5–2 mm),
- Iodine-starch test of leaf sheaths
to determine the appropriate timing and amount of topdressing fertilizer.
The size of the spikelets, which influences 1000-grain weight, is also determined by nutritional conditions during this phase.
Between the end of midseason drainage and the panicle initiation stage, switch to intermittent irrigation to promote healthy root development.
From heading to maturity (the grain-filling period), starch production becomes the primary physiological activity. During this period:
- Leaf growth ceases,
- Leaves begin to senesce, and
- Photosynthetic ability declines.
However, about 70% of the starch required to fill the grains and ensure 1000-grain weight depends on the photosynthesis of upper leaves, including the flag leaf, during this stage.
To enhance photosynthesis:
- Maintain sufficient nitrogen nutrition after heading
- Ensure a high number of green leaves
- Promote good light exposure
- Keep leaf moisture levels high through late stages
Nutrient management should avoid nitrogen deficiency and include silicon application to improve light posture and suppress transpiration. For water management, saturated irrigation should be continued through the late grain filling stage.
References
- Shozo Matsushima: Improvement and Techniques of Rice Cultivation, 1973 (Yoken-do)
- Noboru Murayama: Overcoming the Law of Diminishing Returns, 1982 (Yoken-do)
- Hideo Okajima: Nutritional Physiology of Rice, 1962 (Nobunkyo)
- Yoshiaki Ishizuka & Akira Tanaka: Nutritional Physiology of Paddy Rice, 1966 (Yoken-do)
- Takamine Matsuo: The Complete Study of Rice – Physiology Volume, 1990 (Nobunkyo)
- Takamine Matsuo: The Complete Study of Rice – Morphology Volume, 1990 (Nobunkyo)
- Edited by Nobunkyo: Basics of Rice Cultivation, 1973 (Nobunkyo)
- Edited by Nobunkyo: Understanding Rice Physiology and Cultivation, 2018 (Nobunkyo)
- Yoshio Shimazaki: Illustrated Guide to Rice Cultivation Technology – Growth and Nutrition, 1969 (Agricultural Technology Promotion Association)
- China Agricultural University Press (2009): Current Research on Soil–Crop and Secondary/Micronutrients in China


